
Immunohistochemical analysis of neurotrophin receptor distribution inmouse peripheral sensory ganglia
Neurotrophin receptor distribution
Authors
Abstract
Aim This study aimed to evaluate the immunohistochemical expression patterns of neurotrophin receptors TrkA, TrkB, TrkC, and p75 NTR in the dorsal root ganglia (DRG) of mice, examining potential morphological differences based on age and sex.
Methods Twelve healthy Balb/c mice were used, comprising young (3 weeks) and adult (6 weeks) groups, with equal numbers of males and females (n=3 per group). DRGs were excised from L4–L5 spinal segments and immunofluorescently labeled for TrkA, TrkB, TrkC, and p75 NTR. Confocal microscopy was used for image acquisition, followed by semi-quantitative analysis. Receptor staining patterns, expression intensity, and subcellular localization were compared across groups according to age and sex.
Results TrkA expression was most prominent in the adult male groups, and was lowest in young females. TrkB showed high expression in adult females and young males, whereas other groups exhibited moderate expression levels. TrkC staining was uniformly weak across all groups, with no significant age or sex- related variation. p75 NTR expression peaked in young males and was notably low in female groups. NeuN, a neuron-specific marker, demonstrated uniform nuclear/perinuclear distribution and similar expression across all groups.
Conclusion The distribution of neurotrophin receptors of DRG is modulated by age and sex. Enhanced expression of TrkA and p75 NTR in males and predominant TrkB expression in females suggest sex-specific neurotrophic signaling mechanisms. These findings highlight the importance of gender-based approaches in understanding the etiopathology and treatment of nerve regeneration and sensorial neuropathic conditions.
Keywords
Introduction
Neurotrophins (NTs) are a family of growth factors that regulate important biological processes in the nervous system, particularly neuronal survival, differentiation, axon growth, and synaptic plasticity. These factors include nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), and others. NTs act on cells in the central (CNS) and peripheral nervous system (PNS) as trophic signals originating from target tissues.1,2 NTs act via high-affinity tropomyosin receptor kinase (Trk) family receptors. These receptors include TrkA, TrkB, and TrkC. The low-affinity p75 neurotrophin receptor can bind all neurotrophins and modulate cellular signaling together with Trk receptors.3 Dorsal root ganglia (DRGs) are important structures that house the cell bodies of sensory neurons involved in peripheral transmission and are highly sensitive to neurotrophin signals.4 Small diameter C-type nociceptors and Aδ/Aβ fibers are dependent on neurotrophin signaling for development and regeneration after injury. The NGF–TrkA axis is known to be vital for the survival of sensory neurons. TrkB and TrkC receptors are found in more limited neuronal subpopulations and play important roles in modulating proprioceptive transmission and mechanical sensitivity. Biological variables such as age and sex have been shown to have important effects on neurotrophin expression and receptor distribution.5 A decrease in the expression of Trk receptors and an increase in p75 NTR are observed with aging, which is associated with a decrease in regenerative capacity.6,7,8
This study examines the distribution of TrkA, TrkB, TrkC, and p75 NTR receptors in the L4–L5 dorsal root ganglia of mice and takes into account age and sex variables.9,10 The aim is to elucidate the relationship between neurotrophin receptor expression and biological determinants, contributing to a more comprehensive understanding of Age- and sex- specific neurotrophic support mechanisms in the nervous system. These findings aim to shed light on the pathogenesis of neurodegenerative diseases and aid in the development of age and gender-matched targeted neurotrophin-based treatment strategies.
Materials and Methods
This study was conducted to evaluate the age- and sex-dependent distribution of neurotrophin receptors (TrkA, TrkB, TrkC, and p75 NTR) in mouse dorsal root ganglia (DRG). This analysis focused on the DRG structures that reflect the morphological and molecular properties of the PNS.
Twelve healthy Balb/c mice (approximately 40 g each) were divided into four groups (n=3 per group) by age and sex: young (3 weeks) and adult (6 weeks) males and females. All mice were housed under standard laboratory conditions (12-hour light/dark cycle) with ad libitum access to feed and water.
Following deep ketamine anesthesia, animals were euthanized, and L4–L5 DRGs were meticulously dissected under a stereomicroscope. Excised ganglia were fixed in 4% paraformaldehyde (4°C, 1h), rinsed in PBS, and cryoprotected overnight in 30% sucrose (4°C). Ganglia were then cryo- sectioned (5–30 µm thickness), mounted on poly-L-lysine- coated slides. Tissue sections were blocked for 1h at room temperature (3% BSA, 0.1% Triton X-100, 0.1% sodium azide in PBS), followed by overnight incubation with primary antibodies at 4°C for double immunofluorescence labeling.
(1) TrkA (rabbit polyclonal, Upstate P04629) + TrkB (mouse monoclonal, Becton-Dickinson 554001).
(2) TrkC (goat polyclonal, Upstate 07-226) + p75NTR (rabbit polyclonal, Chemicon AB1554)
Some sections were labelled with NeuN (mouse anti-NeuN; Chemicon MAB377) for neuronal nuclei. Then the sections were incubated for 1 h with appropriate secondary antibodies conjugated with Alexa Fluor® 488, 568, or 647 (Molecular Probes), followed by DAPI-containing mounting medium.
Prepared slides were imaged using a Zeiss LSM 510 META confocal microscope and analyzed with Zeiss Axiovision and ZEN software.
Semi-quantitative evaluation of neurotrophin receptors (TrkA, TrkB, TrkC, p75 NTR) was performed across age and sex groups. Parameters assessed included fluorescence intensity (weak, moderate, strong), staining pattern (granular vs. homogeneous), subcellular localization (perinuclear, cytoplasmic, membrane-associated), association with neuronal diameter (small, medium, large), and involvement of peripheral structures (axonal processes, perineurium, endoneurium). A visual scoring scale (– to +++) was performed by two independent investigators.
Double-labeling used specific polyclonal/monoclonal antibody pairs for TrkA/TrkB and TrkC/p75 NTR. NeuN immunolabeling identified neuronal structures. Expression profiles were comparatively analyzed by age and sex.
Ethical ApprovalThis study was approved by the Animal Care and Use Ethics Committee of the Experimental Medicine Research Center of Selçuk University (Date: 31.05.2010, Decision No: 044).
Statistical AnalysisSemi-quantitative immunofluorescence data were analyzed using non-parametric methods due to the small sample size and ordinal scoring system. Group differences according to age and sex were evaluated using the Kruskal–Wallis test with Dunn’s post hoc test for pairwise comparisons. Data are presented as median (min–max). Interobserver agreement was assessed using Cohen’s kappa coefficient. All statistical analyses were performed using SPSS version 25.0 (IBM Corp., Armonk, NY, USA), and a p<0.05 was considered statistically significant.
Reporting GuidelinesThe study was reported in accordance with STROBE guidelines.
Results
The expression patterns of neurotrophin receptors and NeuN within mouse DRG were assessed comparatively by age and sex. TrkA expression was most prominent in the adult and young male groups (score: 3), whereas female groups demonstrated lower levels (score: 2). TrkB expression peaked in young males and adult females (score: 3), with moderate expression in adult males and young females (score: 2). TrkC exhibited uniformly low expression across all groups (score: 1) without notable variations based on age or sex. p75 NTR showed the highest immunoreactivity in the young male group (score: 3), moderate expression in adult males (score: 2), and weak expression in both female groups (score: 1). NeuN staining was consistently strong across all groups (score: 3), with no observable differences related to age or sex. The comparative immunohistochemical expression levels are shown in Figure 1.
Morphological Distribution PatternsTrkA expression in adult and young males was strong, widespread, and granular within the cytoplasm, with no clear association with neuronal diameter. In the female groups, staining was weaker, sparser, and showed an asymmetric distribution.
TrkB expression was strong and homogeneous in young males and adult females, with prominent staining localized to the plasma membrane and neuronal processes. While TrkB was more concentrated in large-diameter neurons, some groups exhibited localized and asymmetric patterns.
TrkC staining was consistently weak in the ganglion tissues. In young animals, weak expression was detected along axonal processes and the perineurium, whereas in adults, it was mostly confined to the inner plasma membrane surface.
p75 NTR expression was strongest in young males, with a granular and perinuclear localization pattern. Moderate expression was observed in adult males, while both female groups demonstrated markedly weak staining. Notably, neurons adjacent to the perineurium exhibited more intense p75 NTR immunoreactivity.
NeuN expression exhibited uniform nuclear and perinuclear distribution across all groups. No NeuN staining was observed in the neuronal processes, and the signal was diffusely distributed throughout the ganglion.
Representative Confocal ImagingIn adult male mice, TrkA and TrkB co-expression was robust and widely distributed in both the cytoplasm and plasma membrane of the sensory neurons (Figure 2).
In young males, TrkA exhibited moderate granular cytoplasmic localization, whereas TrkB demonstrated strong membrane- bound fluorescence. TrkC and p75 NTR showed weak but distinct staining along peripheral axons and perineurial regions. Triple labeling revealed nuclear co-localization of NeuN in p75 NTR-positive neurons (Figure 2).
In adult females, TrkA and TrkB co-expression was primarily membranous with moderate cytoplasmic signals. TrkC and p75 NTR were co-expressed in specific neuronal subpopulations, particularly around perineuronal structures(Figure 3). In young females, TrkA staining was weak, with a granular cytoplasmic pattern, and TrkB showed moderate membrane-associated expression(Figure 3).
Discussion
This study represents the first systematic evaluation of the immunohistochemical distribution of TrkA, TrkB, TrkC, and p75 NTR receptors in the DRG with respect to age and sex. The findings demonstrate that neurotrophin receptor expression varies significantly depending on both developmental maturation and biological sex.
TrkA is a high-affinity receptor for NGF that is predominantly expressed in small-diameter nociceptive neurons. Approximately 40% of DRG neurons in adult mice are TrkA-positive, making it one of the largest DRG subpopulations.11 Our results showed no age-related differences in TrkA immunoreactivity in the L4– L5 DRGs. This aligns with the existing literature reporting that TrkA labelling remains largely stable in mice aged 3–24 months, and found no significant age-related changes in TrkA expression.12 Even in aged rats (30 months), only a mild reduction in TrkA immunoreactivity has been reported.13,14 This suggests that neurotrophic support in nociceptors is largely preserved during aging.
Our study suggests that TrkA expression is higher in male mice, consistent with previous findings showing more TrkA+ nociceptors in males and more TrkB+/TrkC+ neurons in females.15 This sexual dimorphism may be influenced by estrogen, which can suppress NGF/TrkA signaling. Estrogen decline, such as during menopause, may enhance TrkA activity and increase pain sensitivity.16 These findings indicate that sex hormones modulate nociceptor phenotypes and may underlie sex differences in pain perception. Thus, therapies targeting the NGF-TrkA pathway might need to be adapted based on age and sex.
TrkB, a high-affinity receptor for BDNF and NT-4/5, is typically found in medium- to large-diameter myelinated sensory neurons.17 In our study, TrkB expression showed no significant age- related differences between young and adult mice, consistent with prior reports, though minor declines have been noted in very old animals.15 BDNF-TrkB signaling is known to increase during tissue injury and inflammation, and elevated BDNF levels have been observed in aged DRGs, possibly reflecting enhanced sensory plasticity with age.
Regarding sex differences, TrkB immunoreactivity was slightly higher in females, aligning with neonatal studies that report more TrkB+ neurons in female DRGs. This may suggest a developmental predominance of mechanoreceptive and proprioceptive subtypes in females. Functionally, sex-specific responses have been noted: for example, BDNF elevation increases pain behavior in male but not female mice, indicating possible differences in downstream signaling.18
Transcriptomic studies in adult human DRGs show similar baseline distributions of neuronal subtypes across sexes. However, sex-specific differences may emerge under pathological conditions like chronic pain, where BDNF-TrkB signaling may operate differently in males and females.19 Thus, even subtle variations in TrkB expression could influence sensory sensitivity and pain susceptibility, highlighting the relevance of personalized treatment approaches.
TrkC is the high-affinity receptor for neurotrophin-3 (NT-3) and a distinguishing marker of large-diameter proprioceptive neurons.10 Both rodent and human studies indicate that TrkC defines a distinct population of large-diameter neurons, typically involved in sensing body position and movement.16 Our current findings indicate that TrkC immunoreactivity, similar to TrkA and TrkB, does not undergo significant changes with aging in the DRG.13 This consistency across Trk receptors suggests that fundamental neurotrophic support for various sensory modalities might be largely maintained throughout the adult lifespan, at least in the peripheral nervous system. Regarding sex differences, our study did not identify any substantial sex-specific variations in TrkC expression within the DRGs.14 This differs from TrkA and TrkB, where subtle sex dimorphisms were observed. This absence of marked sexual dimorphism for TrkC may imply that proprioceptive pathways, crucial for motor coordination and spatial awareness, are less subject to sex hormone modulation compared to nociceptive or mechanoreceptive pathways.20 However, further research is warranted to fully elucidate any nuanced sex-specific roles of TrkC under various physiological and pathological conditions. The p75 neurotrophin receptor (p75 NTR) binds all neurotrophins with low affinity and mediates both survival and apoptotic signals, unlike Trk receptors. It is highly expressed in most DRG neurons during early development but decreases significantly postnatally.13 In adults, p75 is mainly found at low levels in small neuron subpopulations and glial cells. Our study showed stronger p75 expression in young mice, consistent with its developmental role, particularly in the transition of non- peptidergic nociceptors from TrkA to Ret dependency.21
Although baseline p75 levels are low in adults, they increase after injury or neurodegeneration. Slight rises have also been noted in aged but healthy DRGs,13 suggesting altered neurotrophin sensitivity with aging. The influence of sex hormones on p75 is not well understood; short-term estrogen may increase its expression, but prolonged exposure might reduce it.16 We found no significant sex differences in baseline p75 levels, though pathological conditions might reveal sex-specific responses, such as greater inflammation in male DRGs after nerve compression. Since elevated p75 is linked to apoptosis and degeneration, it may contribute to sex differences in neurodegenerative vulnerability. Overall, p75 expression decreases with age and shows minimal sex-based variation under normal conditions. Understanding its regulation under stress could aid in developing sex-specific treatments for neuropathic pain and nerve repair.
NeuN is a common marker for mature neurons, and its count reflects the total neuronal population in the DRG. In our study, NeuN+ counts showed no significant differences between young and adult mice, indicating stable sensory neuron populations from postnatal development to adulthood. No sex-based differences were observed either, consistent with transcriptomic studies showing similar neuron subtype distributions in adult male and female DRGs.11,13 However, transient sex differences exist during early development, with female neonates having more neurons and males showing more advanced maturation, differences that appear to even out by adulthood14 Although neuronal number remains stable in adulthood, aging is associated with neuronal size reduction and accumulation of degenerative lipofuscin pigments.16 Both age and sex thus influence not only receptor expression but also neuronal integrity.
Our study is the first to comprehensively analyze TrkA, TrkB, TrkC, p75 NTR, and NeuN expression in mouse L4–L5 DRGs across age and sex. While previous research has mainly examined these factors separately, we show subtle but meaningful sex differences -such as higher TrkA in males and greater TrkB/TrkC in females- highlighting the biological impact of sex hormones and development on sensory neurons.12,19
These findings help explain why pain sensitivity and chronic pain disorders vary by age and sex. For example, increased BDNF with age may underlie heightened pain in older adults, and hormonal modulation of neurotrophin receptors likely influences sex-specific pain perception and nerve repair.16,18 Clinically, this underscores the importance of considering age and sex in treating peripheral neuropathies and pain syndromes, as therapies targeting NGF-TrkA or BDNF-TrkB pathways may have different effects depending on these factors.
Recent studies on human DRGs have revealed sex-specific molecular pathways implicated in neuropathic pain, with elevated IL-1β and TNF expression in male DRGs and increased chemokine and opioid-related gene expression in female DRGs.22 These findings reinforce the need for personalized pain therapy.
Limitations
There were some methodological limitations in the study. First, limited subgroup stratification (n=3 per age-sex group) restricts the statistical power and generalizability of the findings of this study. While semi-quantitative scoring provides valuable morphological insights, it lacks the objectivity of quantitative molecular approaches. Furthermore, the cross-sectional study design precluded longitudinal assessment of neurotrophin receptor dynamics over developmental stages. Potential inter- animal variability in receptor expression, despite controlled housing and genetic homogeneity, could not be excluded. Finally, although the double-labeling approach revealed co- localization patterns, functional validation of receptor activity or downstream signaling was beyond the scope of this study.
Conclusion
This study systematically evaluated the immunohistochemical distribution of TrkA, TrkB, TrkC, and p75 NTR receptors in the dorsal root ganglia (DRG) of mice with respect to age and sex variables. The findings revealed that TrkA and p75 NTR expression were more prominent in the adult male group, whereas TrkB expression was more pronounced in adult females. TrkC expression remained consistently low across all groups, showing no significant age-related increase but exhibiting receptor-specific variations associated with sex.
These results demonstrate that neurotrophin receptor expression is not confined to developmental stages alone but continues to play an active regulatory role in the mature nervous system of the adult mouse. Moreover, sex-based differences in neurotrophic receptor profiles may contribute to variations in neurophysiological outcomes, such as nerve regeneration capacity and chronic pain sensitivity. These findings provide a foundational reference for developing age- and sex-adapted neurotrophin-based therapeutic strategies and enhance our understanding of peripheral nerve pathophysiology.
Declarations
Ethics Declarations
All experimental procedures involving animals were conducted in accordance with institutional and national guidelines for the care and use of laboratory animals and were approved by the relevant ethics committee.
Animal and Human Rights Statement
All procedures performed in this study were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Informed Consent
Not applicable due to the animal study design.
Data Availability
The data supporting the findings of this article are available from the corresponding author upon reasonable request, due to privacy and ethical restrictions. The corresponding author has committed to share the de-identified data with qualified researchers after confirmation of the necessary ethical or institutional approvals. Requests for data access should be directed to bmp.eqco@gmail.com
Conflict of Interest
The authors declare that there is no conflict of interest.
Funding
None.
Author Contributions (CRediT Taxonomy)
Conceptualization: Z.H., E.E.
Methodology: Z.H., E.E.
Formal Analysis: Z.H.
Investigation: Z.H.
Data Curation: Z.H.
Writing – Original Draft Preparation: Z.H.
Writing – Review & Editing: E.E.
Supervision: E.E.
Abbreviations
BDNF: Brain-Derived Neurotrophic Factor
CNS: Central Nervous System
DRG: Dorsal Root Ganglion
NGF: Nerve Growth Factor
NT: Neurotrophin
PNS: Peripheral Nervous System
Trk: Tropomyosin Receptor Kinase
References
-
Peach CJ, Tonello R, Damo E, et al. Neuropilin-1 inhibition suppresses nerve growth factor signaling and nociception in pain models. J Clin Invest. 2024;135(4):e183873.
-
Lorenzini L, Baldassarro VA, Stanzani A, Giardino L. Nerve growth factor: the first molecule of the neurotrophin family. In: Calzà L, Aloe L, Giardino L, eds. Recent advances in NGF and related molecules. Cham, Switzerland: Springer; 2021:3-10.
-
Chaldakov GN, Aloe L, Yanev SG, et al. Trackins (Trk-targeting drugs): a novel therapy for different diseases. Pharmaceuticals (Basel). 2024;17(7):961.
-
Dusan M, Jastrow C, Alyce MM, et al. Differentiation of the 50B11 dorsal ganglion cells into NGF and GDNF responsive nociceptor subtypes. Mol Pain. 2020;16:1744806920970368.
-
Xiong LL, Chen L, Deng IB, Zhou XF, Wang TH. P75 neurotrophin receptor as a therapeutic target for drug development to treat neurological diseases. Eur J Neurosci. 2022;56(8):5299-318.
-
Numakawa T, Odaka H. The role of neurotrophin signaling in age-related cognitive decline and cognitive diseases. Int J Mol Sci. 2022;23(14):7726.
-
Shanks HRC, Chen K, Reiman EM, et al. p75 neurotrophin receptor modulation in mild to moderate Alzheimer disease: a randomized, placebo-controlled phase 2a trial. Nat Med. 2024;30(6):1761-70.
-
Nordvall G, Forsell P, Sandin J. Neurotrophin-targeted therapeutics: a gateway to cognition and more? Drug Discov Today. 2022;27(10):103318.
-
de Assis GG, de Sousa MBC, Murawska-Ciałowicz E. Sex steroids and brain- derived neurotrophic factor interactions in the nervous system: a comprehensive review of scientific data. Int J Mol Sci. 2025;26(6):2532.
-
Singh M, Krishnamoorthy VR, Kim S, Khurana S, LaPorte HM. Brain- derived neuerotrophic factor and related mechanisms that mediate and influence progesterone-induced neuroprotection. Front Endocrinol (Lausanne). 2024;15:1286066.
-
Rostock C, Schrenk-Siemens K, Pohle J, Siemens J. Human vs. mouse nociceptors - similarities and differences. Neuroscience. 2018;387:13-27.
-
Vincent K, Dona CPG, Albert TJ, Dahia CL. Age-related molecular changes in the lumbar dorsal root ganglia of mice: signs of sensitization, and inflammatory response. JOR Spine. 2020;3(4):e1124.
-
Dodge ME, Rahimtula M, Mearow KM. Factors contributing to neurotrophin- independent survival of adult sensory neurons. Brain Res. 2002;953(1-2):144-56.
-
Bergman E, Fundin BT, Ulfhake B. Effects of aging and axotomy on the expression of neurotrophin receptors in primary sensory neurons. J Comp Neurol. 1999;410(3):368-86.
-
Vradenburgh SA, Van Deusen AL, Beachum AN, et al. Sexual dimorphism in the dorsal root ganglia of neonatal mice identified by protein expression profiling with single-cell mass cytometry. Mol Cell Neurosci. 2023;126:103866.
-
Seol SH, Chung G. Estrogen-dependent regulation of transient receptor potential vanilloid 1 (TRPV1) and P2X purinoceptor 3 (P2X3): implication in burning mouth syndrome. J Dent Sci. 2022;17(1):8-13.
-
Haberberger RV, Barry C, Dominguez N, Matusica D. Human dorsal root ganglia. Front Cell Neurosci. 2019;13:271.
-
Hayashi K, Lesnak JB, Plumb AN, et al. Brain-derived neurotrophic factor contributes to activity-induced muscle pain in male but not female mice. Brain Behav Immun. 2024;120:471-87.
-
Tavares-Ferreira D, Shiers S, Ray PR, et al. Spatial transcriptomics reveals unique molecular fingerprints of human nociceptors. Sci Transl Med. 2022;14(632):eabj8186.
-
Barcelon E, Chung S, Lee J, Lee SJ. Sexual dimorphism in the mechanism of pain central sensitization. Cells. 2023;12(16):2028.
-
Chen Z, Donnelly CR, Dominguez B, et al. p75 is required for the establishment of postnatal sensory neuron diversity by potentiating ret signaling. Cell Rep. 2017;21(3):707-20.
-
Ray PR, Shiers S, Caruso JP, et al. RNA profiling of human dorsal root ganglia reveals sex differences in mechanisms promoting neuropathic pain. Brain. 2023;146(2):749-66.
Figures
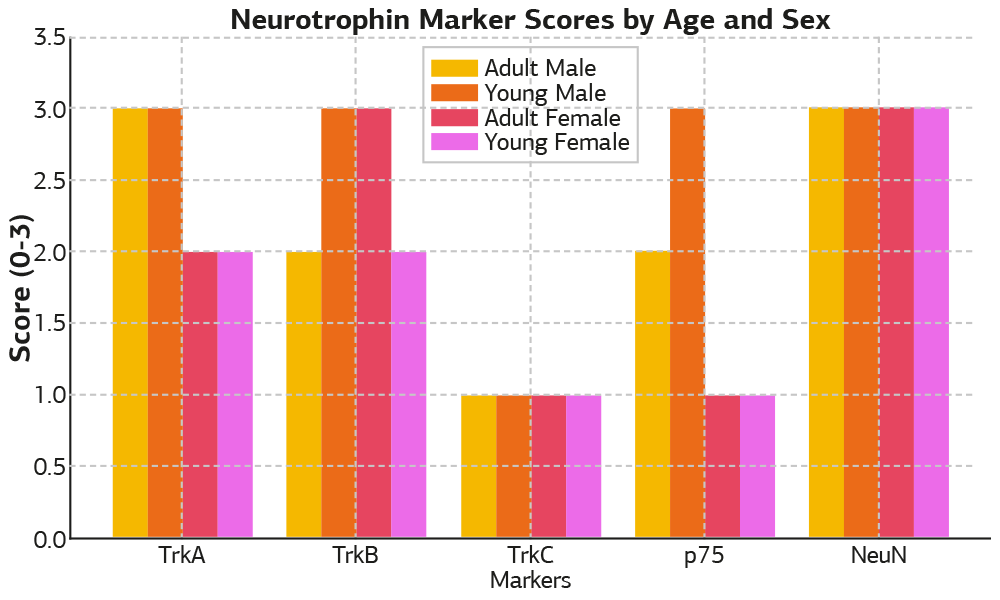
Figure 1. Neurotrophin marker staining scores by age and sex (scale 0–3). Semi-quantitative scoring of neurotrophin receptor expression by age and sex in mouse dorsal root ganglia.
Expression levels of TrkA, TrkB, TrkC, p75, and NeuN were semi- quantitatively scored (0 = none, 1 = weak, 2 = moderate, 3 = strong) in L4–L5 dorsal root ganglia of BALB/c mice. Scores were presented comparatively across four groups: young males, young females, adult males, and adult females
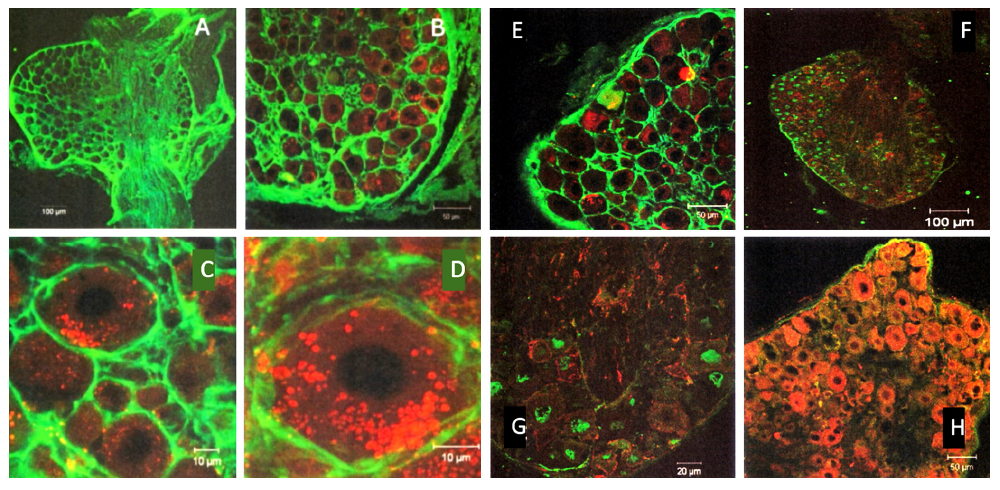
Figure 2. Confocal micrographs of TrkA and TrkB co-expression in adult male dorsal root ganglia (Left 2 columns; A, B, C, D).
Representative confocal images showing the immunoreactivity of TrkA (red, Alexa Fluor® 564) and TrkB (green, Alexa Fluor® 488) in the DRG sections of adult male mice. Double-labeling revealed cytoplasmic and membranous localization patterns in the sensory neurons. Immunofluorescence patterns of neurotrophin receptors in young male dorsal root ganglia. Confocal sections of young male DRG showing (Right 2 columns; E, F, G, H): (E) Co-expression of TrkA (red) and TrkB (green); (F, G) TrkC (green) and p75 NTR (red); (H) Co-labeling of p75 NTR (red) and NeuN (blue). The fluorophores used included Alexa Fluor® 488, 564, and 647. Scale bars were shown in the figures
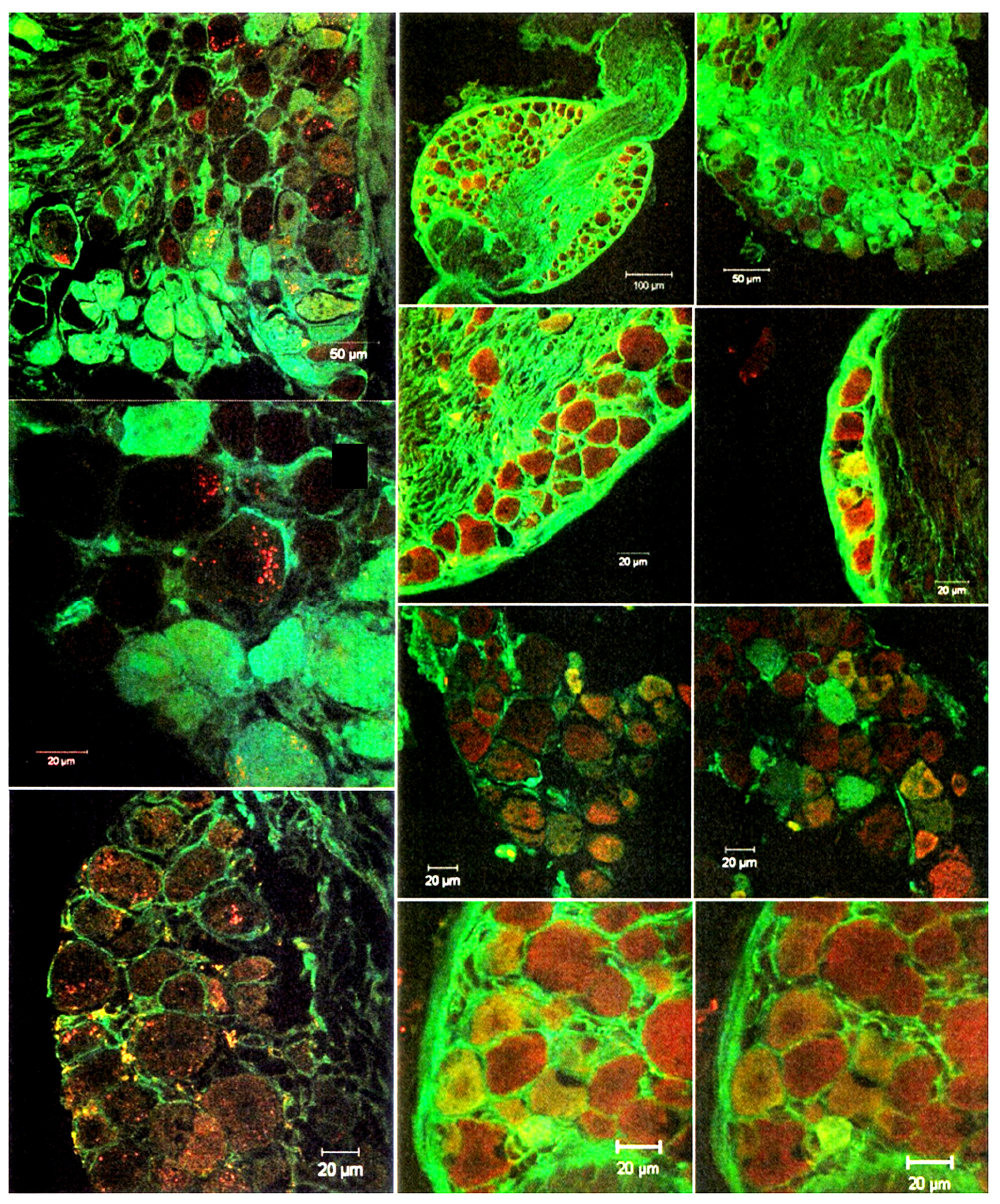
Figure 3
Additional Information
Publisher’s Note
Bayrakol MP remains neutral with regard to jurisdictional and institutional claims.
Rights and Permissions

This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC 4.0). To view a copy of the license, visit https://creativecommons.org/licenses/by-nc/4.0/
About This Article
How to Cite This Article
Zeynep Hayırlıoglu, Ender Erdoğan. Immunohistochemical analysis of neurotrophin receptor distribution inmouse peripheral sensory ganglia. Ann Clin Anal Med 2026;17(5):439-445. doi:10.4328/ACAM.22775
- Received:
- June 16, 2025
- Accepted:
- August 25, 2025
- Published Online:
- October 21, 2025
- Printed:
- May 1, 2026